Structure of Plants
This reading describes what we refer to as the basic structure of a "typical plant." As you go through other readings at this site, you will realize that what we consider "typical" is only a convenient place to start. The example we use for a typical plant is the type of plant with which you probably are most familiar—a flowering plant.
An appropriate essay for you to read as you start learning about the basic structure of plants is "How to Spend a Nice Quiet Evening With a Potato" by Edgar Anderson. It is enjoyable yet very informative.
Basic Structural Plan
A flowering plant consists of an axis divided into two main systems—the root and shoot. The shoot system is composed of a stem, leaves, and flowers. Roots, stems, leaves, and flowers often are referred to as plant organs.
Growth
Roots and shoots grow at specialized regions. Apical meristems occur at the tips of either roots or shoots. Apical meristems account for what is called primary growth, the type of growth responsible for the increase in length of a plant. All flowering plants have primary growth. Examples of apical meristems are apical buds of stems, axillary buds of stems, and tips of roots.
Some flowering plants have growth originating from regions termed lateral or secondary meristems. Growth from lateral meristems is secondary growth. Secondary growth accounts for most of the thickening of plants. Trees, for example, have secondary growth that increases the diameter of the stems, or "tree trunks." The secondary meristem responsible for the increase in diameter of stems is called the vascular cambium. Vascular cambium also occurs in roots of trees.
Stems
Stems generally occur above ground, and are the structures onto which leaves are attached. This latter point is very important for distinguishing stems from roots since some stems occur below ground.
Leaves attach to stems at nodes. The regions between nodes are called internodes. Even when leaves are not attached to a stem one can generally recognize the nodes and internodes. On many stems when leaves are not attached a leaf scar is where the leaf was attached. Small buds (axillary buds) occur at the nodes near where leaves attach to the stem. This is another way of recognizing a node if a leaf is not present.

Primary growth of stems occurs at apical meristems. The apical meristems of stems differ from those of roots: apical meristems of stems are enclosed in an apical bud. In longitudinal view the apical bud contains young leaves (leaf primordia) attached at nodes. The leaf primordia enclose the apical meristem. In winter buds of trees the leaves that will burst out in bright green in the spring are protected by bud scales on the outside.
Further back from the bud, cells differentiate. The differentiated tissues in a young stem are best described when the stem is viewed in cross section. There are two basic patterns of differentiated tissues in stems of flowering plants. We look at other reasons for making distinctions between two types of flowering plants later in the course. For now, however, we just have to know that one type is called monocots, the other dicots.
Dicot stems The epidermis is on the outside of the stem. Toward the interior of the stem just underneath the epidermis is the cortex. The most interior region of the stem is the pith. In the region between the pith and the cortex are several to many vascular bundles arranged in a ring. The phloem is on the side of the bundle facing toward the cortex; the xylem on the side toward the pith.
In plants that increase in diameter as they get older, e.g., trees, the increase in diameter is due to secondary growth. Secondary growth is produced by a lateral or secondary meristem, the vascular cambium. Vascular cambium originates from a group of cells between the xylem and phloem. As the vascular cambium produces new cells, thus new secondary growth, xylem cells are formed to the interior of the vascular cambium, phloem cells are formed to the exterior of the vascular cambium. The xylem is commonly called wood. The bark is everything to the exterior of the vascular cambium; major components of bark include phloem and cork.
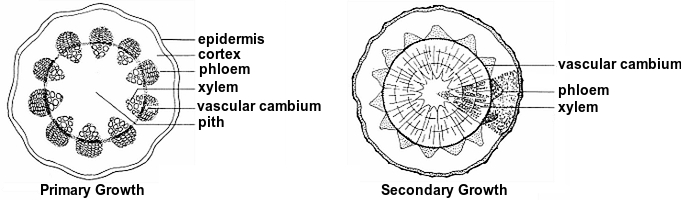
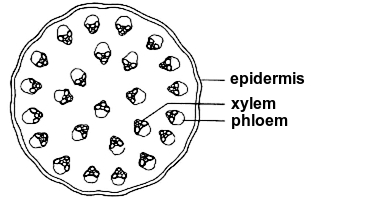
Monocot stems In monocot stems viewed in cross section, the vascular bundles are scattered throughout the stem. Thus, there is no clear separation of tissues around the vascular bundles into cortex and pith. In the vascular bundles of monocots, the phloem is on the side facing toward the epidermis, the xylem toward the center of the stem. Most monocots do not have secondary growth.
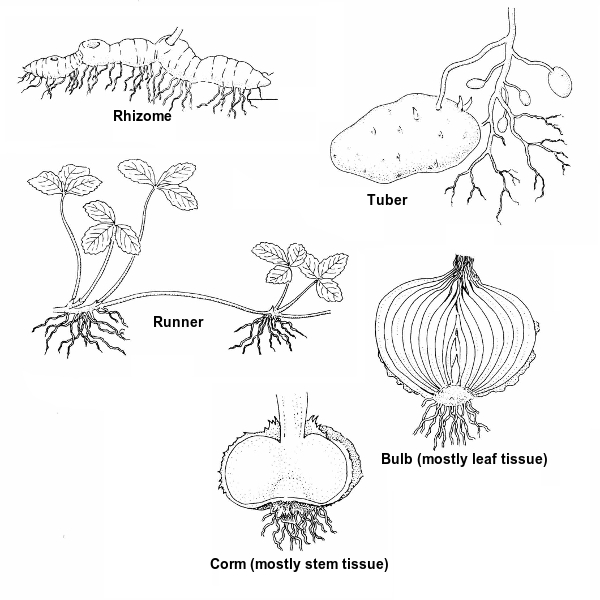
Stem modification Stems of different plants may be in forms other than what we encounter in a "typical" plant. If a stem is underground and horizontal it is called a rhizome. An above ground horizontal stem is a runner, which also is known as a stolon. A swollen tip of an underground stem is a tuber. A vertical underground shoot that is mostly stem tissue is a corm. A vertical underground shoot that is mostly leaf tissue is a bulb. The important part for you to remember here is that if a structure looks like it could be either a stem or a root, look for nodes. Stems have nodes; roots do not!
Roots
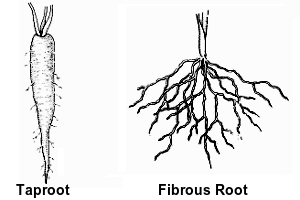
Roots generally grow underground and anchor the plant into the soil. The major function of roots is absorption of water and minerals from the soil. There are two basic types of roots. 1) Taproot has a main, large root that has branch roots coming off of it. A carrot is an example of a taproot. 2) Fibrous roots have many roots, none of which is more prominent than the others. Roots of grasses are examples of fibrous roots.
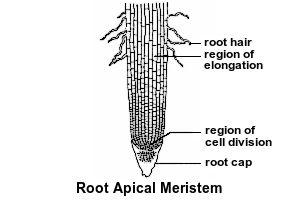
Primary growth of roots occurs at the root apical meristem. In longitudinal view, a root apical meristem has a region of dividing cells, with elongating cells behind them and a root cap in front of them. The root cap is constantly being worn down as the root tip is pushed through the soil by the elongating cells. New root cap cells come from the region of dividing cells. New elongating cells also come from the region of dividing cells.
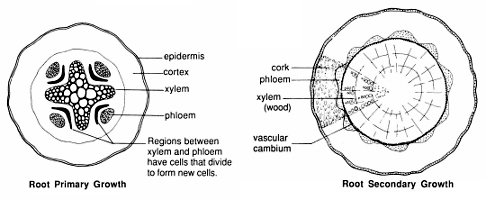
As cells elongate they also begin to differentiate. Some epidermal cells become modified into root hairs, cells that absorb water and minerals from the soil. Cells in the interior of the root differentiate into cells specialized for water and mineral transport (xylem cells); others become specialized into cells that transport sugars. The latter transport cells are termed phloem cells. Xylem and phloem, form a cylinder in the interior of the root known as the stele. The cortex separates the vascular cylinder from the epidermis. As with stems, the patterns of xylem and phloem differ in cross sections of roots of monocots and dicots.
Roots branch. In fibrous systems there are many branch roots, all of which remain about the same size. In taproot systems the main root, the taproot, has smaller roots branched off from it. In either system, however, there is no predictable pattern for the branch roots.
In some plants, e.g., trees, roots increase in diameter and become woody. The growth that increases the diameter of the root is secondary growth. Secondary growth originates at the vascular cambium, a ring of special cells that start dividing between the phloem and xylem. As the vascular cambium produces new cells, cells to the interior of the root differentiate into xylem. Cells to the exterior mostly differentiate into phloem. A mature, woody root is mostly xylem when seen in cross section. The material to the outside of the vascular cambium is the bark.
Leaves
Leaves are attached to stems at nodes. Leaves generally are green because the cells of leaves contain chloroplasts. Chloroplasts are green because of the presence of the green pigment chlorophyll. Chlorophyll is the molecule that converts light energy into chemical energy in the process of photosynthesis.
A leaf is divided into two regions; 1) blade or lamina, and 2) petiole. The petiole is the thin, stick-like structure between the blade and the attachment site at the node. Sometimes at the base of the petiole is a small leaf-like structure called the stipule. If there is no petiole, the leaf is said to be sessile.
Vascular tissue (xylem and phloem) is continuous from the stem, through the petiole, into the blade. Vascular tissue in the blade is seen as so-called veins. The continuation of the vascular tissue from the petiole into the blade is the midrib of the leaf blade. The pattern of the veins is the venation of the leaf. Leaves may have either netted or parallel venation. Leaves of most dicots have netted venation; leaves of most monocots have parallel venation.

Monocot vs. dicot leaves: left is daylily, a monocot; right is mint, a dicot.
The edge of a leaf blade is the margin. Several common variations of margins include smooth (also termed entire), dentate (shaped like teeth), serrate, or lobed.
A simple leaf has one blade attached to the petiole. A compound leaf, however, has several to many little leaves, leaflets (pinnae), in place of a single blade. If the leaflets are attached to one point at the end of the petiole the leaf is palmately compound. If the leaflets are attached along the extension of the petiole, the rachis, in a feather-like arrangement, the leaf is pinnately compound.

Pinnately compound leaf on left; palmately on right.
Leaves can be arranged at the nodes in one of three basic ways. Opposite arrangement is when two leaves are attached at one node on opposite sides of the node. Alternate arrangement is when one leaf is attached to a node, and at the next node one leaf is attached on the other side of the stem. Whorled arrangement has three or more leaves attached at one node.
Flowers
The portion of stem to which a flower is attached is the peduncle. The receptacle is the swollen tip of the peduncle to which the other major flower parts are attached. Sepals occur in the outermost whorl of leaf-like structures, the calyx. The next whorl to the interior is the corolla; the individual leaf-like structures are the petals. If sepals and petals look the same, they are called tepals. To the interior of the petals are the stamens, the pollen-producing structures. Each stamen consists of a thin filament and sac-like structure, the anther, in which pollen is produced. The most interior structure of the flower is the pistil, which is divided into three parts, the stigma, style and ovary. Seeds are produced in the ovary.

External and cut-away view of flower of Lilium sp.
Flowers vary many ways: The most obvious are size and color. Another major variation is the number of parts. In monocots, flower parts occur in threes or multiples of three. For example, there may be three sepals, three petals, six stamens, and one pistil. Dicots generally have flowers with parts in fours or fives, or multiples of four or five.

Monocot vs. dicot flowers: left is spiderwort, a monocot; right is phlox. a dicot.
Symmetry is another major way flowers may vary. Flowers with radial symmetry are actinomorphic. Flowers with bilateral symmetry are zygomorphic.

Floral symmetry: left is actinomorphic flower of spreading evening primrose; right is zygomorphic flower of kudzu.
Flowers that contain all major parts (i.e., sepals, petals, stamens, and pistil) are complete. If any one of the four major parts is missing, then the flower is incomplete. If a flower has both stamens and pistil, that flower is perfect. If a flower does not have either stamens or pistil, then that flower is imperfect. An imperfect flower with stamens but no pistil is a staminate flower, also called by some a "male" flower. An imperfect flower with a pistil but no stamens is a pistillate flower, also called by some a "female" flower.
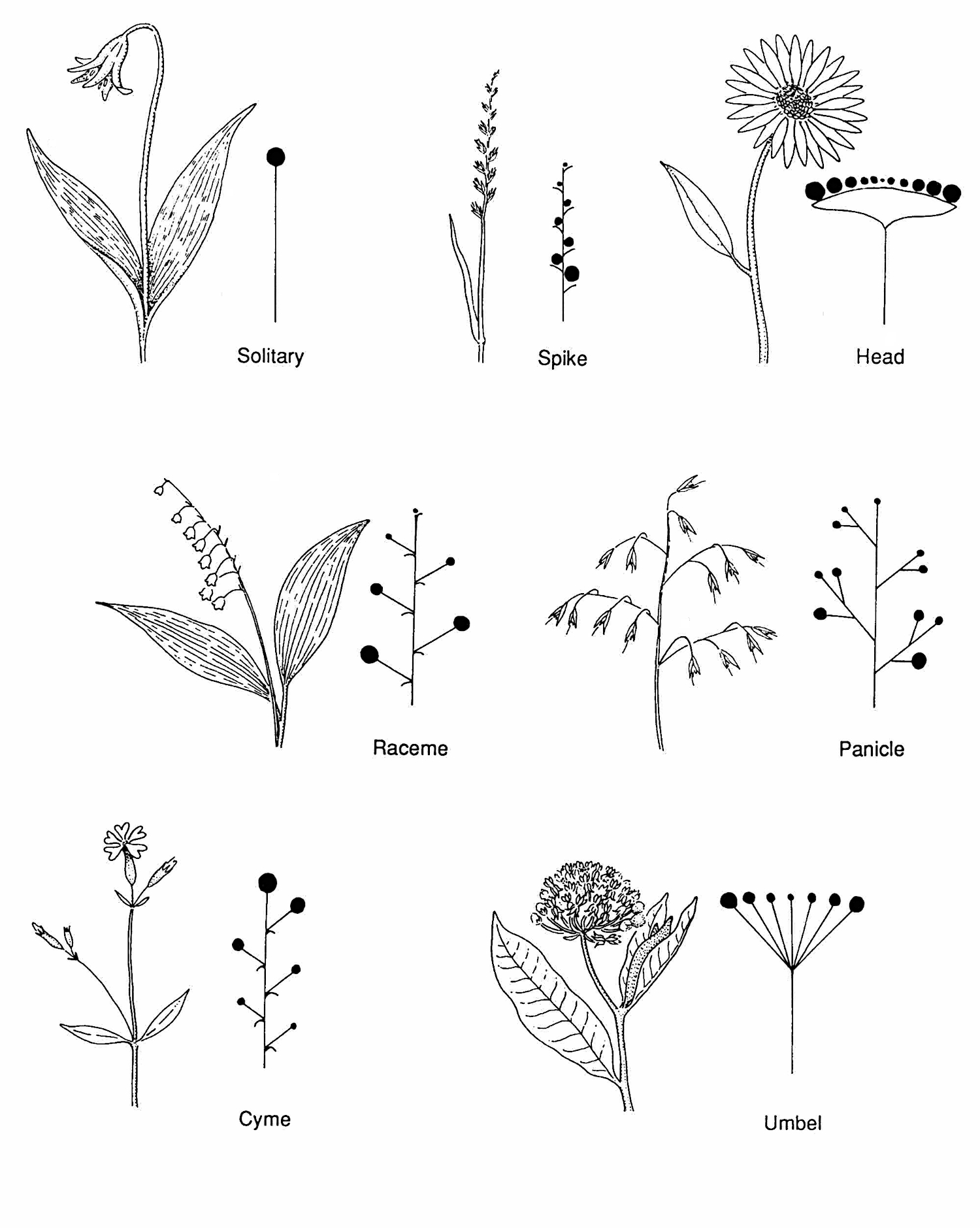
Inflorescences
A group of flowers is an inflorescence. Common inflorescence types are solitary, spike, head (capitulum), raceme, panicle, cyme, and umbel.