The flowering plants (aka angiosperms) are the most common plants on earth at the present time. For this reason most people think of flowering plants when they think of plants in general.
Oak trees and maple trees are flowering plants. Likewise, the grasses are flowering plants even though their flowers are not as showy as those of the great white Trillium, the red bud tree, or the wild crabapple. Some plants have flowers with all the parts (pistil, stamens, petals, and sepals), i.e., the flowers are complete. Other plants have flowers with just pistils, petals and sepals on one individual (imperfect flowers); and flowers with just stamens, petals and sepals on other individuals (also imperfect flowers). Some flowering plants, the annual herbs, take only one growing season to complete their life cycle (go from seed-to-seed). Others such as the oak trees take many years.
There is no one angiosperm that would serve as an example that could represent the entire group. So this discussion of the life cycle we use an idealized plant, one that in principle represents the major features of the group.
The flowering plants that we observe day-in-and-day-out are the sporophytic phase of the life cycle. The two main structures involved in the life cycle and seed formation are the stamens and pistil. Angiosperms, like gymnosperms, are heterosporous, producing two types of spores.
Meiosis within many cells within the anther of the stamens produces many microspores. Microspores are haploid and develop into microgametophytes. Microgametophytes also are called pollen. Up to now in general terms the development of pollen in flowering plants is similar to pollen development of gymnosperms. But pollen of angiosperms is much simpler at maturity than gymnosperm pollen. A mature angiosperm pollen grain consists of three cells: A large cell called the tube cell contains within it two smaller cells, the sperm.
At maturity, the anther splits and pollen is released. Depending on the plant, pollen is then transferred to the pistil either by wind or an animal such as an insect or bird. The transfer of pollen from anther to pistil is pollination.
The pistil consists of stigma, style, and ovary. Pollen lands on the stigma portion of the pistil. We leave the pollen sitting on the stigma and concentrate for the moment on the ovary.
Within the ovary are ovules. This is an important difference between gymnosperms and angiosperms: ovules of angiosperms are enclosed within an ovary! The number of ovules within an ovary varies with species. We are going to jump ahead just a little: the ovary will develop into a fruit; ovules develop into seeds. Think of some fruits and the seed inside and you should get the idea of how ovules are arranged within ovaries. A peach has one seed; only one ovule was within the ovary. A watermelon has many seeds; many ovules were within the ovary. More about fruits later. Let's get back to development within the ovule and how an ovule develops into a seed.
The outer layer of the ovule is the integument. There is an opening at one end of the integument, the micropyle. The interior of the ovule is the megasporangium which contains one cell that goes through meiosis. Only one haploid megaspore results from meiosis.
The megaspore develops into a megagametophyte. There is a big difference, however, between megagametophytes of gymnosperms and angiosperms. The mature megagametophyte of angiosperms is called the embryo sac and consists of seven cells with a total of eight nuclei. The large, central cell contains two nuclei, the polar nuclei. At the micropylar end of the embryo sac is the egg nestled between two cells. Some botanists consider the two cells adjacent to the egg as a very small archegonium.
Remember the pollen we left sitting on top of the stigma? While we were watching the megaspore develop into the embryo sac the pollen germinated. A pollen tube started growing down through the stigma and style, and through the ovary wall. As the tip of the pollen tube grew through the stigma, style, and ovary, the two sperm were carried right behind the growing tip of the pollen tube. We now have found the pollen tube growing right up into the micropyle.
A lot of exciting things happen! First, the pollen tube ruptures. As this occurs the two sperm are discharged into one of the cells next to the egg. One sperm passes through this cell and fuses with the egg and the diploid zygote is formed. We knew that would happen. But the other sperm also passes through the cell and goes into the large central cell. Here the second sperm fuses with the two polar nuclei! This fusion product contains three nuclei, hence three sets of chromosomes. The second fertilization forms a triploid (3n) nucleus called the primary endosperm nucleus. More about the primary endosperm nucleus in a little while.
First let's review what has just happened. One sperm fused with the egg to form the zygote. The second sperm fused with the polar nuclei to form the primary endosperm nucleus. We now have seen something that is unique to flowering plants: double fertilization.
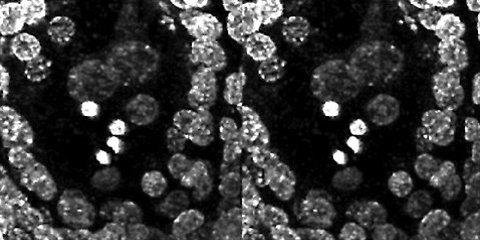
Stereo pair of intact ovule showing the moment of double fertilization in potato. One sperm nucleus is near the two polar nuclei and the other sperm nucleus near the egg nucleus. The other two bright objects are known as "x-bodies," one the remnants of the vegetative (tube) nucleus and the other the nucleus of the degenerated cell that the pollen tube entered.
The confocal scanning laser microscopy images are presented so that the two side-by-side images form a stereo pair for viewing with a standard stereo viewer or map-reading device, or by relaxing and crossing your eyes until the right and left images of each pair fuse into one.
Reference: Braselton, J. P., M. J. Wilkinson, & S. A. Clulow. 1996. Feulgen staining of intact plant tissues for confocal microscopy. Biotech. & Histochem. 71: 84-87.
A seed is a matured ovule. The integument of the ovule develops into the seed coat. The zygote develops into an immature sporophyte, the embryo. The primary endosperm nucleus divides and after many divisions fills the seed around the embryo with a triploid tissue called the endosperm. The mature angiosperm seed consists of old sporophyte (seed coat), a new sporophyte (embryo), and another product of fertilization, the endosperm.
Recall that the ovule is contained within an ovary. While the ovule is maturing, so is the ovary. The ovary matures into a fruit.

Two views of a fruit (legume) of Scotchbroom. In the upper photo, remnants of all floral parts are present. In the lower photo which had strong light shining through the fruit, the unfertilized ovules are small (small arrow) whereas the fertilized ovules (large arrow) will matured into seeds.
We essentially have completed the life cycle of an angiosperm. If the seed is put in the right environment it will germinate. If it is the seed for an annual plant, the seedling will rapidly develop into a mature plant that will flower and produce seeds and fruit within one growing season. Examples of annuals are beans, peas, and corn. Seeds of biennial plants germinate and grow into a fairly mature plant the first year; the plant will not flower and form fruits and seeds, however, until the second year. If the seed is of a perennial plant, the seedling will develop into a larger plant that may take many years to mature before it flowers and produces seeds and fruits.
Let us now turn our attention to some basic variations in seed structure of angiosperms. In other readings the terms monocot and dicot were used to distinguish between the two major groups of flowering plants. Several differences in flower structure, stem structure, and leaf structure have been mentioned. What ultimately distinguishes monocots from dicots, however, is structure of the embryo within the seed.
An embryo is a miniature plant, consisting mainly of an axis with an embryonic shoot and embryonic root. Near the junction of the embryonic root and shoot is attached a leaf-like structure that is called either a seed leaf or cotyledon. Embryos in seeds of monocots have only one cotyledon, hence the designation monocotyledon (one cotyledon). All the flowering plants with seeds that have embryos with a single cotyledon are placed in the Class Monocotyledonae. Another term for the single cotyledon is scutellum.
Two cotyledons are attached to the embryo in the dicots. All the flowering plants with embryos with two cotyledons are placed in the Class Dicotyledonae.
Another major difference generally occurs between seeds of dicots and monocots, but it is not a definitive characteristic. Monocot seeds generally have endosperm present at maturity. The endosperm serves as a food source for the embryo when it starts to grow during germination of the seed. Dicot seeds generally do not have endosperm present. Instead, food from the endosperm was transported into and stored within the two cotyledons during maturation of the seed. The two cotyledons take up most of the bulk of the dicot seed. As the embryo starts to grow upon germination of the seed, food stored in the cotyledons is used for the initial source of energy for the young plant.
In addition to seed structure, there are four other differences between monocots and dicots. (Keep in mind that the definitive difference between the two groups is the number of cotyledons attached to the embryo.)

Monocot vs. dicot flowers: left is spiderwort, a monocot; right is phlox. a dicot.

Monocot vs. dicot leaves: left is daylily, a monocot; right is mint, a dicot.